Identifizierung eines molekularen „Schalters“ für das Wachstum von Blutgefäßen
Forschungsbericht (importiert) 2009 - Max-Planck-Institut für molekulare Biomedizin
Einleitung
Das Blutgefäßsystem ist ein stark verzweigtes Netzwerk aus Arterien, Kapillaren und Venen, das für unsere Gesundheit von zentraler Bedeutung ist. Die kleinsten Verästelungen der röhrenförmigen Blutgefäße durchdringen sämtliche Organe und Gewebe und versorgen alle lebenden Zellen des Körpers mit Sauerstoff und Nährstoffen (Abb. 1). Gefäßerkrankungen, wie Verschlüsse von Blutgefäßen beim Schlaganfall oder sackförmige Gefäßerweiterungen (Aneurysmen), können in Patienten zu schweren gesundheitlichen Schäden führen und zeigen eindrucksvoll, wie wichtig die einwandfreie Funktion von Blutgefäßen ist. Seit Jahrzehnten suchen Forscher nach Möglichkeiten, die Bildung neuer Blutgefäße und damit auch die Reparatur von Organschäden gezielt zu steuern. Andererseits nutzen auch Tumore einwachsende Blutgefäße für ihre eigene Versorgung mit Sauerstoff und Nährstoffen. Das Gefäßnetz im Tumor ermöglicht gleichzeitig die weitere Verteilung von Krebszellen über den Blutstrom und fördert somit die Bildung von Metastasen. In diesen Fällen erscheint es sinnvoll, das Wachstum von Blutgefäßen zu unterdrücken. Somit erfordern unterschiedliche Erkrankungen gezielte Verfahren, mit denen Neuverzweigungen im Gefäßnetzwerk, je nach therapeutischem Ziel, stimuliert oder gehemmt werden können [1; 2].
 grün markiert.")
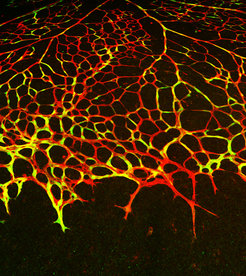
Das Wachstum von Blutgefäßen erfordert eine Serie komplexer molekularer und voneinander abhängiger Prozesse, wie z. B. die koordinierte Erzeugung verschiedener Zelltypen und ihren Zusammenbau zu einem funktionellen vaskulären System. Darüber hinaus muss bei dem Auswachsen neuer Kapillaren gewährleistet sein, dass die Stabilität und Funktionalität der bereits vorhandenen Blutgefäße nicht wesentlich eingeschränkt wird, weil ansonsten die geschwächte Gefäßwand Blutungen und unzureichende Versorgung des angrenzenden Gewebes verursachen kann. Wachstum von Blutgefäßen findet im sich entwickelnden Organismus statt – im Embryo entsteht das Blutgefäßsystem als eines der ersten Organe und gewährleistet so das Wachstum und die normale Entwicklung des Embryos. Doch auch im erwachsenen Organismus behält das Gefäßsystem seine Plastizität und kann bei Bedarf erweitert werden, was beispielsweise durch veränderte Sauerstoff- und Nährstoffbedürfnisse im Gewebe oder durch Regenerationssignale ausgelöst wird.
Die Forscher am MPI für molekulare Biomedizin beschäftigen sich mit dem kardiovaskulären System von Wirbeltieren und der zentralen Fragestellung, wie sich das Blutgefäßsystem während der Entwicklungsprozesse erweitert, in ein Netzwerk aus Arterien, Kapillaren und Venen differenziert und sich präzise in verschiedene Organsysteme einfügt. Ein besseres Verständnis dieser physiologischen Prozesse bildet auch die Grundlage für die therapeutische Beeinflussung von Blutgefäßen bei menschlichen Erkrankungen.
„An- und Aus-Schalter“ auf Endothelzellen – Kontrollpunkte beim Gefäßwachstum
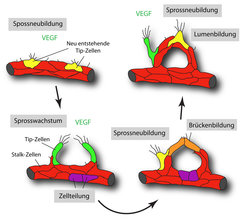
Unzureichende Gefäßversorgung und Sauerstoffmangel führt in den betroffenen Geweben zur Ausschüttung des Wachstumsfaktors VEGF (Vascular Endothelial Growth Factor), der entsprechende Rezeptoren in den Endothelzellen, also in der innersten Zellschicht der Gefäßwand, aktiviert. VEGF löst dabei in diesen Zielzellen sehr unterschiedliche Effekte aus. Manche Endothelzellen verändern ihre Morphologie, werden mobil, entwickeln kleine zelluläre Fortsätze (Filopodien), die sich zur Quelle des VEGF hin orientieren, und bilden die Spitze eines neuen Sprosses. Andere Zellen, die wahrscheinlich weniger stark auf VEGF ansprechen, bleiben zurück oder bilden die Basis des Sprosses. Gleichzeitig vermehren sich die Endothelzellen und bilden somit die für das Auswachsen eines neuen Blutgefäßsprosses erforderliche Zellmasse. Erst die Summe dieser verschiedenen Prozesse, die oft unter dem Überbegriff „Angiogenese“ zusammengefasst werden, ermöglicht ein effizientes Wachstum des Gefäßnetzes (Abb. 2), [1; 2].
 entwickelt zelluläre Fortsätze, wird mobil und invasiv und bildet die Spitze (Tip) eines neuen Sprosses. Andere Endothelzellen bleiben zurück und bilden die Sprossbasis (Stalk). Durch den Kontakt von Tip-Zellen entstehen neue brückenartige Kontakte (orange), die anschließend in neue röhrenförmige Gefäße umgewandelt werden. Gleichzeitig erzeugt Zellteilung (lila) zusätzliche Endothelzellen.")
Ob und wie die Gefäßröhren oder einzelne Endothelzellen auf VEGF reagieren, hängt von einem molekularen Schalter ab, der jüngst durch die Arbeit unserer Abteilung identifiziert worden ist. Die zentrale Komponente dieses Schalters ist ein Rezeptor mit dem Namen „Notch“, der sich auf der Oberfläche von Endothelzellen befindet. Die Interaktionspartner (Liganden) von Notch sind ebenfalls Zelloberflächenproteine der Genfamilien Jagged und Delta, sodass Bindung und Signaltransduktion durch Zell-Zell-Kontakt ausgelöst werden [3]. Diese Moleküle spielen wesentliche Rollen in vielen verschiedenen Organismen und biologischen Prozessen. Mehrere Mitglieder beider Ligandenfamilien finden sich auch im Endothel, wobei insbesondere die Liganden Delta-like 4 (Dll4) und Jagged1 eine essentielle Rolle bei der Angiogenese spielen. Die Bindung zwischen Dll4 und Notch unterdrückt die Expression von VEGF-Rezeptoren in Endothelzellen und hemmt deren Sprossung und Vermehrung. Dll4 bewirkt somit, dass der Schalter für Blutgefäßwachstum in die „Aus“-Position umgestellt wird, was auch bereits in früheren Arbeiten gezeigt worden ist. Vollkommen neu ist hingegen die Erkenntnis, dass der Ligand Jagged1 diese Dll4-Notch-Interaktionen entscheidend stören und damit Angiogenese aktivieren kann, d. h. den Schalter für Gefäßwachstum in die „An“-Position umstellen kann [4].
Jagged1 – der überraschende Gegenspieler
Die Eigenschaft von Jagged1 als molekularer Gegenspieler ist zunächst sehr überraschend, da der Ligand in vielen biologischen Prozessen die Aktivierung von Notch-Rezeptoren bewirkt. Die Arbeit der Gruppe um Ralf H. Adams hat nun aufgedeckt, dass Zucker-Modifizierungen an Notch entscheiden, wie die verschiedenen Liganden mit dem Rezeptor interagieren. Ist Notch in Endothelzellen nicht modifiziert, so wirken Jagged1 und Dll4 in gleicher Weise als schwache Aktivatoren. Dagegen potenziert die Modifizierung von Notch die Aktivierung durch Dll4, während Jagged1 nur noch als schwacher und somit unzureichender Aktivator fungiert [4]. Jagged1 kann dabei auch Interaktionen zwischen Dll4 und Notch vermindern und dadurch die Notch- Signaltransduktion hemmen. Die Folge: Viele Endothelzellen präsentieren VEGF-Rezeptoren an der Zelloberfläche, können VEGF binden und bewirken so das Wachstum neuer Gefäßsprosse (Abb. 3).

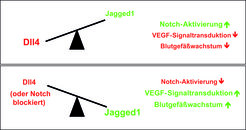
Die Balance beider Liganden entscheidet also, wie stark Notch aktiviert wird. Interessanterweise findet sich Dll4 besonders stark auf Endothelzellen an der Spitze von Sprossen, wodurch die Expression von VEGF-Rezeptoren auf den Nachbarzellen reduziert und somit die Reaktion auf VEGF verringert wird. Diese Nachbarzellen an der Basis eines Sprosses besitzen wiederum weniger Dll4 und sehr viel Jagged1, wodurch die Aktivierung von Notch-Rezeptoren in der Sprossspitze vermieden wird. Dadurch spricht die Zelle in der Spitze weiterhin stark auf VEGF an und die resultierenden positiven Signale fördern das weitere Auswachsen des Gefäßsprosses. Dieser molekulare Schaltmechanismus wirkt sowohl auf zellulärer Ebene als auch im gesamten Gefäß. Auf zellulärer Ebene kontrolliert die Balance zwischen Dll4 und Jagged1 die Entscheidung, welche Endothelzellen sprossen oder im Gefäß verbleiben werden (Abb. 3). Überwiegen Dll4-Notch-Interaktionen aber im gesamten Gefäß, so werden VEGF-induzierte Spross- und Zellteilungsprozesse komplett gehemmt. Zu der umgekehrten Reaktion, d. h. starker Sprossung und Teilung, kommt es, wenn der Notch-Rezeptor blockiert wird oder Zellen überwiegend Jagged1 präsentieren [4].
Hoffnung auf neue Therapieansätze
Die Entdeckung eines molekularen Steuermechanismus des Gefäßwachstums durch die Proteine Dll4 und Jagged1 mit gegensätzlicher Funktion könnte neue Möglichkeiten zur Behandlung von Gefäß- und Krebserkrankungen eröffnen. Bereits seit Jahren wird die Hemmung des Wachstumsfaktors VEGF bei der Behandlung von Krebspatienten und bestimmten Augenerkrankungen therapeutisch eingesetzt [5]. Allerdings ist die Therapie nur bei einem Teil der Patienten erfolgreich und zeigt zudem eine Reihe unerwünschter Nebenwirkungen, welche den therapeutischen Einsatz deutlich einschränken. Die Blockierung von Jagged1, beispielsweise durch spezifisch bindende Antikörper, könnte ebenfalls bestehende Gefäße stabilisieren und die Bildung neuer endothelialer Sprosse in solchen Patienten unterdrücken. Umgekehrt könnte eine vorübergehende Hemmung von Dll4 eingesetzt werden, um die Bildung neuer Gefäße anzuregen. Dies wäre nach Organtransplantationen oder generell bei Regenerationsprozessen sinnvoll.
Die Aufklärung der Funktion von Jagged1 könnte somit völlig neue Therapieansätze möglich machen. Allerdings muss die Wirksamkeit und die Verträglichkeit derartiger Verfahren zunächst gründlich in verschiedenen Tiermodellen geprüft werden, um die Grundlage für die Entwicklung zukünftiger Therapeutika beim Menschen zu schaffen. Eine Komplikation ist bereits vorhersehbar, da der Rezeptor Notch und seine beiden Liganden Dll4 und Jagged1 auch in anderen Zelltypen und Organen wichtige Aufgaben haben. Es ist eine anspruchsvolle Herausforderung, die therapeutische Wirkung nur auf die gewünschten Zelltypen zu beschränken.