Umprogrammierung von Körperzellen in pluripotente Stammzellen
Forschungsbericht (importiert) 2005 - Max-Planck-Institut für molekulare Biomedizin
Einleitung
Zelltherapien wird zur Behandlung degenerativer Erkrankungen gegenwärtig ein viel versprechendes Potenzial eingeräumt, wobei defekte oder nur noch eingeschränkt funktionsfähige Zellen durch frische ersetzt werden sollen. Unter der Vielzahl der zu lösenden Probleme bei diesen Therapieansätzen stechen einige besonders heraus: i. Häufig gibt es nur sehr limitierte Möglichkeiten für die Zellisolation aus Spenderorganen (z.B. Insulin-produzierende Zellen der Bauchspeicheldrüse, Nervenzellen, Herzmuskelzellen). ii. Selbst wenn geeignete Zellen aus Spenderorganen prinzipiell isoliert werden können, stehen sie oft nicht in ausreichendem Maß zur Verfügung (z.B. Leberzellen). iii. Differenzierte Zellen aus dem erwachsenen Körper (somatische Zellen) lassen sich meist nur eingeschränkt vermehren, sodass es schwierig ist, genügend Zellen für eine Therapie zu bekommen. iv. Um Abstoßungsreaktionen zu vermeiden, wären Patienten-eigene Zellen oder Zellen von Spendern mit sehr ähnlichen Gewebsmerkmalen (Major Histocompability Complexes, MHC) wünschenswert.
Gerade im Hinblick auf die eingeschränkte Verfügbarkeit geeigneter Zellen bieten Stammzellen, die sich selbst erneuern, eine wichtige Perspektive. Nach anfänglicher Euphorie wird den leicht zu isolierenden adulten Stammzellen des Knochenmarks jedoch international nur noch ein eingeschränktes therapeutisches Potenzial eingeräumt; nicht zuletzt deshalb, weil der Mechanismus der vermuteten Transdifferenzierung, also der Umprägung von Zellen des einen Gewebetyps in die eines anderen, häufig nicht bestätigt werden konnte.
Möchte man aber Zellen in der Kulturschale als Transplantate für bestimmte Erkrankungen vordifferenzieren, gewinnen embryonale Stammzellen (ES) eine zunehmende Bedeutung. Diese stammen aus der inneren Zellmasse einer Blastozyste, also einer befruchteten Eizelle noch vor ihrer Einnistung in die Gebärmutter und sind noch pluripotent, was bedeutet, dass sie noch zu allen Zelltypen des Organismus ausdifferenzieren können. Um zum einen Verfahren ohne Embryonen entwickeln zu können und zudem die molekularbiologische Prozesse besser untersuchen zu können, wäre es hilfreich, wenn embryonale Stammzellen an Stelle von Eizellen verwendet werden könnten. Im Gegensatz zu Eizellen können nämlich ES-Zellen in großer Zahl eingesetzt werden, was biochemische Untersuchungen erleichtert, etwa wenn wichtige Proteine isoliert werden sollen. Außerdem lassen sich durch die Verwendung von ES-Zellen selbst seltene Transformationsereignisse leicht selektiv vermehren, was mit Eizellen nicht möglich ist. Schließlich wären Verfahren mit ES-Zellen eher akzeptabe für all jene, die ethische Bedenken gegenüber dem Ableiten neuer ES-Zellen oder dem „therapeutischen Klonen“ haben. Dabei wäre gerade letzteres Verfahren besonders geeignet, um MHC-kompatible oder Patienten-identische embryonale Stammzellen zu etablieren. Solche Stammzellen wären vorteilhaft, um Abstoßungsreaktionen transplantierter Zellen zu vermeiden und um Krankheitsmodelle in der Kulturschale zu entwickeln.
Ein Verfahren der Forschergruppe um Hans Schöler besteht darin, bereits existierende embryonale Stammzellen mit somatischen Zellen zu fusionieren, die diese so umprogrammieren („verjüngen“) können, dass sie selbst wieder einen pluripotenten Phänotyp aufweisen. Mit diesem Verfahren könnte man sowohl Patienten-spezifische Stammzellen gewinnen als auch eine „Stammzellbank“ mit wenigen hundert Zelllinien aufbauen, die den Hauptbedarf an Gewebe-kompatiblen Zellen abdecken würde.
Umprogrammierung von Körperzellen in pluripotente Stammzellen
Somatische Zellen können in vitro de-differenziert werden, indem sie mit pluripotenten Zellen fusioniert werden und somit Gene aktiviert werden, die in adulten Stammzellen nicht aktiv sind. Ein charakteristisches Gen für die Pluripotenz von Stammzellen ist Oct4, ein Transkriptionsfaktor, der die Expression und damit die Aktivierung einer Vielzahl von Genen reguliert. Die Etablierung einer transgenen Maus, die ein grün-fluoreszierendes Protein (GFP) unter Kontrolle des Oct4-Promotors exprimiert, macht es möglich, die Pluripotenz von Zellen sichtbar zu machen: Oct4-positive Zellen fluoreszieren grün und sind ein erster wichtiger Hinweis, dass sie pluripotent sind.
In Vorarbeiten konnte die Arbeitsgruppe bereits zeigen, dass Oct4 bzw. ein Fusionsprotein aus Oct4 und green fluorescent protein (GFP) als Pluripotenzmarker nach dem Transfer eines somatischen Kerns in eine entkernte Eizelle (Oozyte) reaktiviert wurde [1; 2]. In den hier zusammengefassten Studien wurde nun untersucht, ob das entkernte Zytoplasma von ES-Zellen ebenfalls das Oct4-GFP-Transgen in somatischen Zellen einer Maus reaktivieren kann oder ob Komponenten des Zellkerns notwendig sind. Hierfür wurden neonatale Nervenzellen, so genannte Neurosphere Cells (NSC) einer Oct4-GFP-Maus gewonnen und mit ES-Karyoplasten bzw. ES-Zytoplasten fusioniert (Abb. 1). Die Ergebnisse dieser Versuche zeigen, dass die mittels Dichtezentrifugation isolierten Karyoplasten die Oct4- Expression induzieren konnten, die verbleibenden Zytoplasten diese Fähigkeit jedoch nicht besitzen. Weitere Untersuchungen wurden mit ES-Zellen durchgeführt, die mit Mitomycin C behandelt waren. In diesen ist die DNA-Replikation und somit die Zellteilung blockiert, wobei sie dennoch 5-Azacytidin-behandelte neonatale Nervenzellen umprogrammieren. Daraus kann geschlossen werden, dass die Fähigkeit zur Reaktivierung von Oct4 in den ES-Karyoplasten liegt und dass diese Genreaktivierung unabhängig ist von DNA-Replikation und Zellteilung [3].
 mit Nervenzellen (Neurosphere Cells, NSC) das grün-fluoreszierend markierte Pluripotenzgen Oct4 aktivierte (A, B). Die Fusion von Zytoplasma mit den Nervenzellen führt hingegen nicht zur Umprogrammierung und somit nicht zur Aktivierung der Fluoreszenz (C, D).")

Bringen neonatale Zellen einen Stammzellcharakter mit, den ausdifferenzierte somatische Zellen nicht haben?
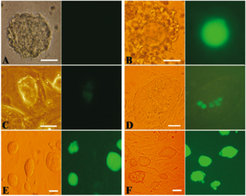
Neurosphere Cells sind in vitro kultivierte neonatale Nervenzellen, die sich in Neurone und Astrozyten ausdifferenzieren können, weswegen sie noch einen eingeschränkten, „bipotenten“ Stammzellcharakter besitzen. Um die Möglichkeit auszuschließen, dass nur dieser „Stammzellvorsprung“ die Umprogrammierung zu pluripotenten Zellen möglich machte, wiederholten die Wissenschaftler die Experimente mit vollständig ausdifferenzierten Zellen. Hierfür wurden Kumuluszellen (Cumulus-Cells, CC) aus Follikeln von Oct4-GFP-Mäusen verwendet, mit ES-Zellen fusioniert und mit NSC-ES-Fusionen verglichen (Abb. 2). Abbildung 2B zeigt die Oct4-negativen Kumuluszellen, welche die grün-fluoreszierende Eizelle umgeben. Das Anschalten des Pluripotenz-Markers Oct4 wird durch die grüne Fluoreszenz erstmals nach ca. 45 Stunden sichtbar (Abb. 2C und D), nach Subkultivierung (Abb. 2E und F) bilden sich schließlich homogene grün-fluoreszierende Zell-Kolonien aus unprogrammierten somatischen Zellen (NSC und CC).
 und Kumuluszellen (CC) vor und nach Fusion mit embryonalen Stammzellen (ES). NSC (A) exprimieren den Pluripotenz-Marker Oct4 nicht und zeigen somit auch keine grüne Fluoreszenz des Oct4-GFP-Transgens. Kumuluszellen (B) sind differenzierte somatische Zellen, die die Eizelle umgeben. Sie zeigen ebenfalls keine grüne Fluoreszenz, wohl aber die von ihnen umgebene Eizelle, die als Keimzelle Oct4-positiv ist. 45 Stunden nach Fusion der NSC und CC mit ES zeigen sich erste umprogrammierte, grün-fluoreszierende Zellen (C, D), die nach weiterer Kultivierung homogen grün-fluoreszierende Kolonien bilden (E, F).
Aus: Jeong Tae Do, Hans R. Schöler: Comparison of neurosphere cells with cumulus cells after fusion with embryonic stem cells: reprogramming potential. Nachdruck mit freundlicher Genehmigung aus Reproduction, Fertility and Development 2005;17:143-149")
Doppelter Chromosomensatz – normale Differenzierung ?
Die zu pluripotenten Stammzellen umprogrammierten somatischen Zellen besitzen durch die Fusion mit den ES-Zellen einen doppelten Chromosomensatz. Dennoch lassen erste Experimente darauf schließen, dass sie sich in Zellarten aller drei Keimblätter differenzieren lassen. Zumindest GluR6 als ektodermaler Marker, HNF3β und α-Fötoprotein als endodermale Marker sowie βH1 und α-cardiac myosin als mesodermale Marker konnten nachgewiesen werden [4]. Trotzdem scheint es notwendig, eine Segregation in Zellen mit normalem Chromosomensatz zu erreichen. Die aktuellen Fragestellungen beinhalten daher: Lassen sich die ursprünglichen Chromosomensätze wieder trennen? Besteht eine Art nukleäres Gedächtnis des Chromatins für seine Ursprungszelle und auch für deren Spezialisierung? Wenn dem so wäre, ließen sich beispielsweise Herzmuskelzellen für Therapien ischämischer Herzerkrankungen einfacher ableiten, wenn für die Umprogrammierung bereits Muskelzellen eingesetzt würden.